Tajemství v oblacích - heliamfory
Roku 1839 objevili bratři Schomburgkové na jihoamerické hoře Roraimě masožravou rostlinu Heliamphora nutans. V té době byl již znám příbuzný rod Sarracenia (viz Živa 1982) a roku 1841 byla poznána ještě Darlingtonia. Dodnes si přírodovědci lámou hlavu otázkou, jaké jsou vývojové souvislosti mezi těmito třemi rody, tvořícími čeleď Sarraceniaceae a vlastně i celý řád Sarraceniales. Na tomto zdánlivě jednoduchém problému, co z čeho se vyvinulo aneb "co bylo dříve", ztroskotávají i moderní taxonomové. Jeho vysvětlení by přitom určitě bylo významným přínosem ke znalostem florogeneze Amerik.
Přírodovědci mají svoje srovnávací studium ztížené tím, že sběr heliamfor na jejich obtížně dostupných nalezištích je skutečným dobrodružstvím. Získání potřebných květů, semen a z hlediska vývojového tolik důležitých semenáčků pomocí kultury se pokládá za mimořádný úspěch. Heliamfory jsou tak ještě dnes tajemstvím obestřené rostliny.
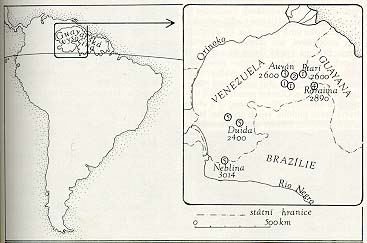
Rozšíření rodu Heliamphora. Velmi blízké lokality jsou značeny společnou kruhovou značkou s číselným označením druhu: 1 - H. heterodoxa, 2 - H. ionasi, 3 - H. minor, 4 - H. nutans, 5 - H. tatei. Obrázky kreslila R. Novotná
V čem je problém
Dnešní místa výskytu jednotlivých rodů čeledi Sarraceniaceae jsou zcela izolovaná. Rod Heliamphora se přitom nachází v jiné květenné říši nežli Sarracenia a Darlingtonia. Heliamfory se také liší od bylin z obou ostatních rodů souborem znaků pokládaných za primitivní, vývojově staré. Je to dřevnatý kmen u H. tatei, u všech druhů květenství pouze jednoobalných květů (oproti jednotlivým a dvouobalným květům ostatních rodů), chybění nektarových žláz ve květech, jednodušší pestík z neustáleného počtu 3-4 plodolistů, láčky nejvíce prozrazující svůj vznik svinutím původně plochého listu aj. Zde se nabízela lákavá hypotéza, že heliamfory jsou nejpůvodnější, takže jsou vývojovými předchůdkyněmi odvozenějších rodů Darlingtonia a Sarracenia. Logické bylo i to, že právě heliamfory se udržují v geohistoricky nejméně proměnlivé tropické zóně a k tomu na neobyčejně starých geologických útvarech.

Heliamphora nutans, juvenilní stadium. Měřítkem je květník o průměru 8 cm
Do této hypotézy ovšem vůbec nezapadá počet chromozómů v buněčných jádrech: Heliamphora 2n = 42, Darlingtonia 2n = 30, Sarracenia 2n=26. Heliamfory by jako nejpůvodnější měly mít počet naopak nejnižší. Existuje také několik dalších rysů naznačujících svéráznost heliamfor: zvláštní vidličnaté chlupy na vnějším povrchu láček i na pestíku, ostatním rodům zcela nepodobná zonace povrchu uvnitř láček, zvláštní typ tyčinek i ve více rysech jiný pestík apod. Heliamfory jsou zřejmě od neznámého společného vývojového základu všech tří rodů řazených dnes do čeledi Sarraceniaceae také značně vzdálené. Podle nejnovějších názorů by měly s ohledem na přirozenost systému dokonce patřit do samostatné čeledi Heliamphoraceae. V literatuře se vyskytuje i názor, že vývojové centrum dosavadní široce pojaté čeledi Sarraceniaceae neleží v Jižní Americe, ale spíše v Severní Americe.
Jaké jsou
Botanici až příliš samozřejmě pokládají kornoutovitě svinuté a po straně jen dole srostlé listy heliamfor za vývojově zaostalý typ láček. Pouhý dotyk s masívně stavěnou a přitom křehkou a lámavou pastí kterékoli heliamfory však přírodovědce intuitivně varuje před takovým unáhleným závěrem. Zde si příroda dala neobyčejně záležet a dala vzniknout solidně stavěnému a ve srovnání s rody Sarracenia a Darlingtonia neméně složitému lapacímu aparátu, šikovně přizpůsobenému zvláštním ekologickým podmínkám tropických vysokohor.
Před vertikálními i horizontálními srážkami rostlina skrývá jen kapku nektaru, důležitou k vábení hmyzu. Ta visí v miskovitě vyklenutém přívěsku, schýleném nad kornoutovitým ústím láčky. Kořist se k nektaru dostane jen po přistání na vnitřním okraji kornoutu, kde je ovšem tuhé a dolů sčesané husté ochlupení.* (* Je zajímavé, že heliamfory vytvářejí také formy s kornoutem víceméně olysalým (H. tatei i. macdonaldae, H. heterodoxa f. glabra). Hmyz zde nachází špatnou oporu, a proto brzy skončí v tekutině obsažené v hojném množství v láčce. Trávicí zóna, tvořící dolní polovinu láčky, je nahoře hladká a lysá, dole při dně je oděna dosti řídkými, dlouhými, dolů sčesanými štětinami. Produkce vlastních rostlinných exoenzymů je u heliamfor potlačena a trávení obstarává symbiotické mikrobiální společenstvo, žijící v tekutině uvnitř láček.
Vystoupavé až vzpřímené láčky heliamfor jsou nahoře široce otevřené, takže do nich prší. Živiny uvolněné z těl kořisti mohou být přijaty stěnami láčky jen částečně a významný zbytek je odplavován s přebytečnou vodou. Ta je odváděna místem, kde jsou okraje horního kornoutu nad trávicí zónou volné. Nad touto odtokovou štěrbinou mohou být okraje kornoutu srostlé i volné. výtok je nasměrován mezi dvě adaxiální lišty (křídla), jež tekutinu s živinami přivádějí až do centra kořenového systému rostliny. Bohatě větvený a silně vyvinutý kořenový systém heliamfor je snad adaptací na využití takového "hnojení".
Stavba květů je rovněž ovlivněna vysokohorskými podmínkami. Obal nicích květů stříškovitě chrání pohlavní orgány před vertikálními i horizontálními srážkami. Všechny části jsou stavěny bytelně a jsou připraveny pro hromotlucké návštěvníky. Těmi jsou, jako všude jinde na horách, zpravidla čmeláci. Jedině oni mohou, podle posledních výzkumů, mohutným vířením svých křídel uvolnit pyl z prašníků.
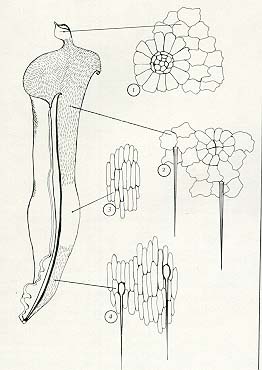

Vlevo proříznutá láčka druhu Heliamphora heterodoxa s patrnou zonací vnitřního povrchu: 1- nektarující přívěsek, 2 - ochlupený zadržovací kornout, 3 - hladká část trávicí zóny (normálně naplněná vodou), 4 - dolní část trávicí zóny s chlupy ukotvenými lahvicovitou bází. V pravé části obrázku mikroskopické zvětšeniny pokožky, v zónách 1 a 2 s hektarovými žlázami, obklopenými paprsčitými buňkami. Vpravo květ druhu Heliamphora minor.
Heliamfory jsou proterogynické, pestík je schopný sprášení až o týden dříve než uzrají tyčinky. Tím je preferováno cizosprášení; v kultuře však byla prokázána možnost umělého samosprášení. Postupuje se při tom tak, že se pyl uvolní z ještě nefunkčního prašníku jeho zlomením. Po oplození květ neopadává, jen dostane zelenavou barvu. Semena vysetá ihned po uzrání na povrch nastříhaného živého rašeliníku klíčí téměř všechna. Semenáčky mají nejprve 2 ploché děložní lístky a pak následuje první láčka. Olistění semenáčků (i nejmenších odnoží starších rostlin) se u všech druhů velmi podobá. Jejich juvenilní láčky jsou trubicovité, s široce ohrnutým obústím a bez miskovitého přívěsku popsaného u láček dospělých rostlin. (Různolistost během vývinu se vyskytuje i u rodů Sarracenia a Darlingtonia.)
Jednotlivé druhy heliamfor se různí svými rozměry i životní formou. Heliamphora tatei var. tatei běžně vytváří až přes metr vysoké kmínky s vrcholovým chocholem láček, a roste-li v křovinách jako opěrná liána, mohou se věkovité exempláře větvit a dosahovat výšky 4 m. Heliamphora ionasi, druh s nejrobustnějšími láčkami, dosahujícími výšky 50 cm, tvoří přízemní růžice. Heliamphora minor s nejmenšími láčkami, vysokými max. 12 cm, roste v kobercích ze stěsnaných růžic, o ploše asi 1 m2
Naleziště
Heliamfory se vyskytují výhradně na "tabulových" či "stolových" horách ve venezuelské části Guayanské vysočiny. Tyto zvláštní geomorfologické útvary s náhorními plošinami jsou nejodolnějšími pozůstatky přes miliardu let staré souše z mocné vrstvy červenohnědých pískovců, jež byla rozrušena a rozčleněna říční erozí a větráním. Ve východní části oblasti výskytu heliamfor se vžilo označení "tepui", což znamená v řeči tamnějších Indiánů kmene Pemon "hora". Jinde se v mapách označují portugalsky slovem "cerro".
Našinec si jako stolovou horu automaticky vybaví cosi jako Říp, anebo tzv. "svědecký vrch" Sovici na Roudnicku. Zde je proto třeba učinit poznámku o rozměrech popisovaných lokalit heliamfor, což jsou hory obrovité a děsivě členité. Nadmořské výšky "stolových hor" jsou běžně nad 2 000 m, ale dosahují i kolem 3 000 m. Kolmá nepřístupná úbočí, někdy členěná ve stupně, jsou od úpatí k vrcholu vysoká kilometr, ale i přes 1,5 km. Plató mívá rozlohu až několik set kilometrů čtverečních. Jeho povrch může být parovinný, ale i rozerodovaný v propasti a bludiště nejroztodivnějších zaobleně členěných skalisek. Ponurou náladu pro výzkumníka a především životní prostředí svérázných biocenóz zde vytváří na tropy studené, mrholivé, deštivé a vysloveně mlhavé podnebí. Opustíme nyní geografický popis, jenž by mohl přejít k ponorným řekám a přečetným vodopádům včetně 979 m souvisle padajícího Salto Angel, a vrátíme se k botanice.
Vysoko na vrcholech "stolových hor" se nacházejí rostlinné formace velmi odlišné od tropického deštného lesa nižších poloh i od Gran Sabany (Velké savany) v prostoru mezi Auyán - tepui a Roraimou. Tyto vegetační rozdíly jsou způsobeny hlavně 6- až 10-stupňovým rozdílem průměrných teplot mezi úpatími a náhorními planinami "stolových hor", kde je nejběžnější teplota kolem 14 °C. Rozdíl průměrné roční teploty 6 až 10 °C je tak značný, že na severní polokouli se takto liší arktoalpínské tundry od listnatých lesů.
Buď lesní, nebo savanový charakter vegetace relativně nízkých poloh pod "stolovými horami" je potom určován kvalitou substrátu a propustností podloží pro vodu. Jisté je, že vzdálenosti mezi "stolovými horami", okupované pralesy a savanou, fungují po geohistoricky dlouhou dobu (asi od pliocénu) jako bariéra bránící šíření vysokohorských druhů z lokality na lokalitu. "Stolové hory" proto mají své vlastní druhy, poddruhy a variety rostlin, čili endemity. Tabulka s přehledem druhů heliamfor ukazuje, že takovými endemity jedné nebo několika těsně sousedících hor jsou i ony.
V posledních letech se stalo módou navštěvovat "stolové hory" vrtulníky s cílem dosáhnout v co nejkratším čase co největšího počtu lokalit a získat tak co největší sortiment heliamfor, bublinatek, genliseí, rosnatek i dalších rostlin. Letecké propojení nalezišť může narušit funkci zmíněné bariéry, a sice transportem semen i jiných diaspor v bahnité půdě ulpívající na podvozcích letadel i v podrážkách bot posádek. Venezuelská vláda umožňuje zatím činnost ne právě nejzodpovědnějším výzkumníkům a botanickým prospektorům.
Vegetace "stolových hor" je ovlivněna nejen velkou nadmořskou výškou a velkou vlhkostí klimatu, ale i zvláštními půdními podmínkami. Projevuje se totiž extrémní chudost matečné horniny na živiny, mnohde nepatrná hloubka půdy na tvrdé skále, v místech konkávních tvarů podloží potom hromadění rašeliny. Rostlinstvo se na základě struktury označuje jako savana. Její součástí jsou porosty nevysokých stromů až křovin, například Bonnetia roraimae (čel. Bonnetiaceae, dříve Theaceae), Weinmannia crenata, W venezuelensis (čel. Cunoniaceae blízká čel. Saxifragaceae), Ilex retusa a četných jiných. Časté jsou zde i keřovitě až stromkovitě rostoucí broméliovité rostliny z rodů Connellia, Navia a Brocchinia ( B. hechtioides a B. tatei), ale vyskytují se i stromovité kapradiny rodů Cyathea a Alsophila, i žebrovnice Blechnum schomburgkii (importována do Botanické zahrady Liberec). Dřeviny jsou střídány plochami nízké, často nezapojené bylinné vegetace.

Listová růžice druhu Heliamphora minor
Heliamfory se vyskytují v polostínu mezi dřevinami i na otevřených plochách, vždy však na mokřadech. Jejich průvodními druhy bývají rašeliníky (Sphagnum magellanicum a S. tenellum rostou i u nás) a další mechorosty, erikoidní keříky z rodu Cyrilla (čel. Cyrillaceae, derivát čel. Celastraceae zastoupené v české květeně rodem Evonymus) a četné jednoděložné byliny, například z rodů Xyris, Orecanthe a Abolboda (čel. Xyridaceae), Paepalanthus a Syngonanthus (čel. Eriocaulaceae), Stegolepis (čel. Rapateaceae), Tofieldia (čel. Liliaceae), Maxillaria (čel. Orchidaceae) i vysokohorské druhy masožravých rostlin Drosera roraimae, D. esmeraldae aj., Utricularia quelchii, U. humboldtii aj. a Brocchinia reducta (čel. Bromeliaceae). Na pustém skalním podkladu se mokřadní vegetace udržuje jen roztroušeně ve vododržných bochníkovitých shlucích, tzv. konsorciích, obsahujících kolem 20 druhů. Vedle mechorostů zde dominují tvrdolisté keříky a tuhé růžicovité byliny. Jejich xeromorfní vzhled by mohl být vysvětlen stejně jako u keříčků arktoalpínských rašelinišť. Někteří ekologové jej chápou jako adaptaci na obzvláštní nedostatek živin, tzv. "hladové formy" - peinomorfózy. Jiní se domnívají, že jde o důsledek špatné propustnosti vody, prosycené organickými makromolekulami z humusu, do cévního systému rostlin. Mokřadní tvrdolisté keříky se skutečně podobají těm ze středomořských či australských suchopárů.
Pěstování
Rostliny z tropických horských až vysokohorských mlžných poloh jsou vždy obtížně pěstovatelné a mají ekologické požadavky splnitelné nejlépe pomocí klimatronu. V letním, u nás příliš teplém období je třeba chladit ovzduší a navodit přitom jeho maximální vlhkost. V zimě je v případě heliamfor nutné také prodloužení osvitu na 12 hodin.
Naštěstí je možné počítat s určitou tolerancí vůči vyšší průměrné denní teplotě než je obvyklá na přirozených stanovištích. Téměř stálá oblačnost a mlhavost na "stolových horách" navíc ovlivnila světelné nároky tak, že heliamfory se v kultuře chovají jako heliosciofyty, takže je možné v létě stínit a chránit je tak před přehřátím. Technické zařízení pro heliamfory může být díky tomu i nedokonalé a značně jednoduché. Musí však zajišťovat v zimě teploty 10 - 17 °C a v létě co nejdelší úsek dne s teplotami pod 20 °C. Několikahodinový vzrůst teplot až na 27 °C v době oslunění rostliny dobře snesou, dlouhodobější působení teplot nad 20 °C však zastavuje růst. Vzdušná vlhkost má být stabilně 80-100 %, přičemž prospívá časté mlžení na listy. Světelně nejlépe vyhovuje východní expozice.
Zařízení, v němž jsem vypěstoval prvé dva získané druhy heliamfor z juvenilního stadia až do květu, mělo jako základ lepenou vitrínu o objemu 150 1, otevíratelnou ze strany. Na stropním skle ležely 2 zářivky po 20 W, ovládané automaticky. Na dně byla novodurová mísa s roštem pro umístění rostlin v hliněných květináčích. Do vody, shromažďující se po zálivce v míse, ústil vývod z malého kompresoru, určeného k vhánění studeného vzduchu nasávaného z obyčejné chladničky. Větrání probíhalo jen malou štěrbinou na horní straně uzávěru vitríny.
Jako vyhovující substrát se osvědčuje živý rašeliník, vsazený do květníku na drenážní vrstvu střepů. Rašeliník má být denně prolévaný měkkou nebo destilovanou vodou. Špatný odtok vody rostlinám v kultuře velmi vadí. Je to zvláštní, protože podle nedávných detailních pozorování různých výzkumníků v přírodě heliamfory zpravidla nekoření přímo v rašelinících, ač s nimi rostou ve společenstvech. často se vyskytují i tam, kde voda v půdě stagnuje. Ekologové však dobře vědí, že pozorování z přírody a z umělých podmínek je možné na vzájem vztahovat jen velmi obezřetně. Přesazování se provádí v únoru a březnu a tehdy lze také provádět množení dělením trsů. Hlavní doba růstu a doba květu je v našich podmínkách na jaře. Příliš dlouhé dny v létě, ve spojitosti s příliš vysokými teplotami, obvykle způsobují útlum růstu. Také v zimě je v kultuře patrný útlum, který je pravděpodobně výrazem přirozené bioperiodicity. Krajina "stolových hor" je totiž oblastí letních dešťů a tropická "zima" bývá srážkově chudším obdobím, i když stále mlhavým. Prakticky bylo odzkoušeno, že heliamfory jsou schopny přežívat i krátkodobé vyschnutí substrátu. Omezení zálivky v zimě se však nedoporučuje, bylo by zbytečné.

Kvetoucí Heliamphora heterodoxa. Snímky M. Studničky
Praktickou pomocí pro pěstitele může být ještě přehled různých pozorovaných poruch vývinu a jejich příčin: Okraje láček se v zimě nepřirozeně silně vyklánějí ven nedostatek světla; okraje většiny láček jsou odumřelé, hnědé - nízká vzdušná vlhkost; v zimě odumřela většina láček, ale střed rostliny je živý - podchlazení až namrznutí; úplné zastavení růstu v létě - vysoká průměrná teplota; rostlina odumírá od středu hniloba navozená původně přehřátím, nebo stagnující vodou v substrátu; rostliny mají normální vzhled, avšak nenabývají na síle - stopy vápníku ve vodě jsou příliš vysoké.
Pro informaci lze uvést také současný sortiment heliamfor v Botanické zahradě Liberec: H. heterodoxa, H. ionasi, H. minor, H. nutans a dosud taxonomicky neurčený druh z úplně nového naleziště. Rostlina byla darována německými objeviteli v roce 1989 a zaslána letadlem přímo z Caracasu. Dosud je v juvenilním stádiu, avšak robustní stavba i nápadně rudé zabarvení láček dávají naději, že půjde o ještě neznámý atraktivní druh
Dosud platně popsané druhy a variety heliamfor
| Druh (varieta) | Velikost láček (cm) | Rok objevu | Naleziště | Vertikální rozšíření (m. n. m.) |
| H. heterodoxa Steyerm. | 20 - 30 | 1951 | Serra do sol, Ptari-tepui, Uei-tepui, Chimantá-tepui | 1 220-2 450 |
| H. heterodoxa var. exappendiculata Maguire et Steyerm | 20 - 30 | 1953 | Churi-tepui, Chimantá-tepui, Torono-tepui, Abacapá-tepui, Aprada-tepui | 1 000-3 000 |
| H. ionasi Maguire | 40 - 50 | 1952 | Ilu-tepui | 2 600 |
| H. minor Gleason | 10 - 12 | 1939 | Chimantá-tepui, Apacará-tepui, Auyán-tepui, Churi-tepui, Abacapá-tepui | 1 865-2 300 |
| H. nutans Benth. | 15 - 30 | 1840 | Roraima | 1 600-2 700 |
| H. tatei Gleason | 40 - 50 | 1928 | Cerro Duida, Cerro Huachamacari | 1 600-2 165 |
| H. tatei var. neblinae (Maguire) Steyerm. | 15 - 25 | 1953 | Cerro de la Neblina | 1 800-2 800 |
Copyright © Miloslav Studnička, 1991